北大・東北大・東大・名大・京大・阪大・九大
卒業生のためのアカデミック・コミュニティ・クラブ
腸内細菌と老化
光岡 知足
(東京大学教授・理化学研究所主任研究員)
No.786(平成2年1月)号
今日私がお話をする演題は〝腸内細菌と老化〟という題でございまして、最初に腸内細菌と老化がどうして関係してきたのかの背景を申し上げ、それから本題に移りたいと思います。
日本は高齢化社会を迎え、世界一の長寿国、男性は七十六歳、女性が八十一歳と平均寿命が長くなりましたことは皆さんもよくご存じのとおりだと思います。そしてこの長寿の方が多くなったことと、腸内細菌とは切っても切れない関係にあるということになるわけです。
長寿の人が多くなりますと、自分は何で死ぬのかということをどなたも気になさると思います。これの一番参考になるのが、死亡因の統計でございまして、明治三十八年の統計では第一位が結核でした。ところが終戦後になりますと、抗生物質が開発され、栄養状態も徐々によくなって結核で亡くなる方、特に若い方では全くいなくなった。現在では抵抗力がなくなったお年寄りの方が結核にかかって亡くなるということはございますけれども、結核で亡くなるということはもうほとんど考えなくてもいいというぐらいになってきたわけです。それでは何の原因で亡くなるかといいますと、昭和四十年、五十年ぐらいまで第一位は脳卒中でした。日本人は脳卒中が非常に多い国民で、日本人の脳血管が弱いということがよく言われました。その次がガンで、第三位が心臓病でした。戦後になりましてからは、いま申しましたように脳卒中、ガン、心臓病という順位は、ずっと変わらないでまいりました。しかし、昭和四十五年頃からこの順序に変化がみられはじめました。一つは脳卒中がだんだん減り始めたということです。もう一つはガンで死ぬけれども、そのガンができる部位が変わってきたということです。日本人は先進国にしては胃ガンが抜群に多い国民でした。ところが、欧米を見ますと、欧米は胃ガンではあまり死なない。それに対して大腸ガン、乳ガンがきわめて多い。先進国としてこういう両極端になっているということが二十年前くらいでしたか、ガン学会における一つの大きな話題でした。そして現在はこのガンの順位、胃ガンが多いということがだんだん崩れてきたことになります。そしてその死亡の原因のうち脳卒中が急激に減りまして、順位からしますと三番目、つまりガン、心臓病、脳卒中の順に変わりました。どうしてこんなに早く変わったのかと言いますと、おそらく日本の経済高度成長と切っても切れない関係があると思います。それは一つに食生活が豊かになってきたということによるようです。いままでは伝統的日本型食事を食べている日本人に、脳卒中が多くて胃ガンも多かったと言えるのではないかと思います。いわばお米を主食にして、漬物を食べ、味噌汁、そして干物ぐらいで済ませた食事、これを少し栄養学的な言葉で言いますと、低蛋白、低脂肪、そして高食塩、高炭水化物、このような食事パターンになると思います。この低蛋白、低脂肪、高食塩、高炭水化物、これがおそらく脳卒中と胃ガンの発生を高めていたと言えるようです。ところが、この二十年ぐらい前から急速にその食生活が欧米化してきました。つまり低蛋白、低脂肪という傾向が、高蛋白、高脂肪、そして食塩はかなり減り、低炭水化物の傾向になってきました。もう一つは食物繊維、いわゆる繊維質を摂らなくなってきていることです。この傾向はアメリカ、ヨーロッパでも同じでございます。こういう傾向に移ってきたということが、おそらく日本人の死亡因がここ二十年間に大腸ガン、乳ガンが多くなってきたことに関係します。またこれは必ずしも食生活とは関係ないのですが、肺ガンも多くなってきました。事実、胃ガンで死ぬ人は少なくなり、これに加えて膵臓ガンや肝臓ガンがふえてきているということがその大きな変化と言えるかと思います。
そして日本人は世界一の長寿国となったのですが、ここに来てどちらかというと心臓病で死にやすくなった。あるいは大腸ガンとか乳ガン、こういう病気で死にやすくなったというふうに変わってきたことでございます。ちなみに、三人に二人、大体六四%の方がガン、心臓病、脳卒中のいずれかで亡くなるわけです。私はいつも感ずるのですが、毎日、新聞に出る有名人の死亡欄を見ていますと、大体そのような率でガン、あるいは心臓病、そして脳卒中で亡くなっているようです。そうなりますと、いま申しました病気に気をつけることが長寿のための秘訣になるのではなかろうかと、私は考えるわけです。
ところで、このような病気、特にガンが大腸ガン型になってきたということ。その大腸ガン、乳ガンが腸内細菌とかかわり合いがあることがわかってきました。つまり老化を防ぐには腸内細菌が重要だということでございます。
さて、私がこの腸内細菌の研究を始めたのは、昭和二十八年、大学院に入ったときにすぐに与えられたテーマであります。これは学術会議の前の会長をされておりました越智勇一先生から授かったもので、私はそれをありがたくちょうだいしてやったわけですが、その当時は、腸内細菌の研究などにはだれも見向きもしませんでした。つまり腸内にいるのは大腸菌と、わかり切ったように考えられていました。ところが、越智先生は「鶏の下痢症を解明する一つの基礎として、腸内細菌がわかっていないとだめだから」といってくださったテーマだったのです。そのときから私は腸内細菌の研究に取りかかりました。そのとき最初に考えついたことは、どうして腸内細菌が大腸菌だけなんだろうかということでした。糞便を直接顕微鏡で見たときと自分が培養してみたときと菌数が全く違うのです。自分の糞便を顕微鏡で見ますと、ものすごいバクテリアの数がありますけれども、培養してみますと、それの千分の一、一万分の一しか培養ができないのです。当時医学の教科書を見ますと、腸内にはたくさんのバクテリアがいるけれども、それはほとんど死んでいる、死菌である、生きているバクテリアのうちでは大腸菌が多いのだというふうに書いてあるのです。ところが、私が顕微鏡で見たのは細菌も一種類じゃない、きわめてたくさんの種類があるわけです。そこで培養法をちょっと改良しましたら、それだけでも培養できる細菌数は十倍、二十倍というふうに変わってまいりました。私はこれは培養法がおかしいのじゃないかということに気がつきました。そこでマスター・コースの学生のときに培養法の改良に従事したわけです。いまでも鮮烈に印象に残っているのは、自分の糞便を培養してみたら、大腸菌はたくさんある培養できた細菌の数にして百分の一以下にしかすぎず、そのほかの種類の細菌が驚くほど多くありました。そこで、どうしてこのような細菌が培養できないのか、自分のお腹だけがおかしいのかと思って、ほかのひとの糞便をいただいて培養してみますと、同じなのです。そのときに一つ特徴あるコロニーの細菌が培養されました。それがいまで言うビフィズス菌だったわけです。当時、私の腸内にビフィズス菌がたくさんいると言いますと、一般の細菌学者は、「それは何か別のものを見ているのでしょう」ということで相手にしてくれませんでした。しかし、私はそのとき信念を持って、あれはビフィズス菌以外にないと思ったわけです。当時、ビフィズス菌は赤ちゃんの菌で、おとなになったら大腸菌に変わるというのが常識でした。しかし、私がつかんだ菌はまさしく赤ちゃんの菌とは違う種類のビフィズス菌だったのです。しかしすぐにはそれがビフィズス菌であることを科学的に証明することができなかったのです。それを分類してみるよう越智先生に言われたお陰で、私はいままでビフィズス菌を研究し続けることができました。ふりかえってみますとそれは、そういうテーマをいただき今日まで研究させてくれた理化学研究所、あるいは東京大学の研究環境が非常によかったということでありまして、あらためて先生方にお礼を申し上げたいと思います。そしてそういう鮮烈な印象がいかに大切なことか、また既成概念や固定観念にとらわれないことがやはり新しい学問を切り開くのにどうしても重要ではないかと、痛感するわけです。
分類学との出会については、私が出ました成蹊高校に、植物の先生として講師で来られていたのが、当時東京大学助教授の前川文夫先生で、後に東京大学の名誉教授になられましたけれども、この前川先生に教えていただいたのです。皆さんはおそらく分類学はおもしろくない嫌な学問とおっしゃるかもしれませんが、植物分類を非常におもしろく、系統発生の道をたどって、なぜこう分類しなくてはならないかを、前川先生から教えていただいたそのことがあったからこそ、このビフィズス菌を分類するように言われたときに、喜んで研究をはじめることが出来たのです。そして、次の十年後、二十年後にそのときの感激が活力になり、また高校のときに教わった前川先生の教えが生きて、自分の一生の仕事にすることができました。少なくとも現在私が、この腸内細菌の学問を開拓することができ、しかも今日皆さんにこれがいかに人の健康にとって大切かということをお話できることを感謝するとともに本当に幸せだと思っております。
では本題に入りまして、この腸内細菌の研究でまず私がびっくりしたことは、大腸菌ではなくて、その百倍も多い菌があるということ、そしてその中にビフィズス菌がいるということでした。そのたくさんの菌を調べてみますと、それまであった考え方と全く違う考え方に立ち至ったわけです。これが実際にわかり発表しましたのが、それから十年ぐらい後でした。まず、人の腸内容を培養してみますと、十二指腸あたりはきわめてバクテリアは少ない。ところが大腸には、百種類のバクテリアがおりまして、その一種類が大腸菌とお考えいただきますと九十九種類はほかの菌で占められています。そして図1のように、大腸では百兆個ものバクテリアを抱えています。百兆個といいますと、毎日排泄される糞便の三分の一ぐらいの体積を占めています。昔はその三分の一を占めているのが死菌であると言われていましたが、それはほとんどすべて生きています。そして、その菌は百種類もおります。つまり皆さんのお腹の中にいつも百種類ものバクテリアを飼っているということです。そして健康であるからには百種類のバクテリアのバランスがかなり一定しております。その中には、善い菌と悪い菌がありまして、そのバランスもいつも一定しております。その中には健康であるからにはビフィズス菌が頑張っております。そのビフィズス菌は赤ちゃんの場合ですと、皆さんの腸の中よりもはるかに多いのです。赤ちゃんの糞便をごらんになった方は多いと思いますが、黄色っぽい色をしていて糞便のにおいがしない。あれはビフィズス菌がつくっている物質で甘酸っぱいにおいがします。ビフィズス菌は主として乳酸と酢酸をつくり、そのほかの悪いにおいのもの、つまり腐敗産物はつくらないわけです。これが赤ちゃんの健康を守っているのです。ところが、赤ちゃんが離乳食になり、おとなに近いものを食べるようになりますと、おとな型の腸内細菌のバランスになってしまいます。昔はビフィズス菌は赤ちゃん時代を過ぎるとなくなると言われていたのですが、実は一〇%から二〇%ぐらいは持っているわけです。私はそのビフィズス菌は善玉の代表だと思います。そして、そのビフィズス菌は赤ちゃんの菌とは違うおとな型のビフィズス菌です。このおとな型のビフィズス菌は不健康な状態になると減ったり、いなくなったりする。もう一つは、ストレスを受けても減ったり、いなくなったりします。
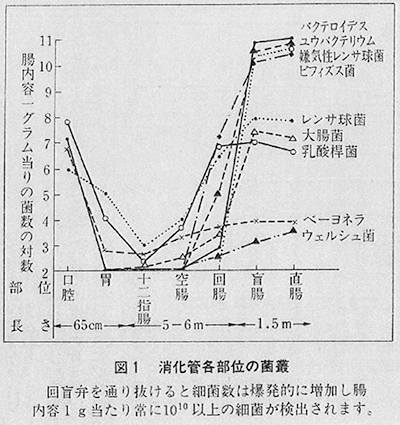
いまから十数年前に、ソ連で宇宙飛行船が打ち上げられたときにこの研究がされました。宇宙飛行中の飛行士のビフィズス菌が減るということがわかりました。すなわち、ストレスを受けますと、ビフィズス菌が少なくなるのです。それから抗生物質を投与したときに真っ先になくなるのはビフィズス菌です。ですから、体の状態が悪くなったときにビフィズス菌が同じように少なくなると考えてよろしいのじゃないかと思います。
腸内細菌のパターン化をしてみますと、赤ちゃん、幼児、成人――成人というのを一応五十数歳までとして、六十歳以上を老人というふうに分けますと、図2にありますような変化がわかってきました。これがきわめて重要なことだったわけです。すなわち、赤ちゃんではビフィズス菌がいっぱいですが、離乳期になりますと、ビフィズス菌が減って一〇%ぐらいになって、一方、糞便のにおいをつけるバクテロイデスとか、ユウバクテリウムとか、嫌気性レンサ球菌が優勢になる。困ったことにお年寄りになりますとビフィズス菌が減ったり、なくなったりします。大体六十歳以上の方を標準にとりますと、十人中三人がビフィズス菌を持っていない。私はこれを一つの老化現象と申し上げております。腸内細菌も老化すると老化現象を起こし、ビフィズス菌がなくなったり少なくなったりします。それに反して悪玉菌と言われるウェルシュ菌がふえてくるのです。この菌が実は非常に臭い便をつくる。普通若い人では大体二人に一人しかこの菌を持っていません。また持っていても取るに足らない程の数しかありません。それがお年寄りになるとふえてくるわけです。その結果腸内の環境はきわめて汚い、悪い環境になり、腐敗が強くなってきます。ですから、このごろ自分の糞便が臭くなってきた、どうしたらいいだろうと思うときには、ビフィズス菌が減って悪玉菌がふえた現象が起こっているのです。つまり臭いということは腐敗物質が多くなっている状態です。そして、腐敗物質は老化を早めるとか、あるいはガンを早めかねません。そのような菌がお年寄りになりますと、どうしてもふえてきます。そこで私は、その現象は単なる腸内の環境が悪くなったと言って放っておけない。若い時代のようには戻りませんけれども、何とかして若い時代に戻すような工夫をしなければならない。その工夫が食生活であり、心の問題ではないかと思うわけです。
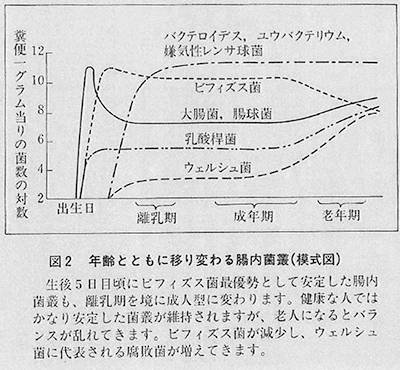
次に、この悪い菌がふえた状態というのはどういう状態かについて説明します。先ほど大腸ガンは高蛋白、高脂肪が関係していると言いましたが、その蛋白質と脂肪を、腸内の特に大腸で待ち構えている悪玉菌がえさにして悪い物質をつくっているわけです。アミンとか、ニトロソアミン、あるいはインドール、スカトール、フェノール、また二次胆汁酸などの悪い物質をつくってしまうのです。そのような物質が、何年後かあるいは何十年後かにガンを引き起こしたり老化を早めたりしかねないということです。もちろん遺伝的にガンになりやすい体質の人は、ほかの人よりも早くガンになることは間違いないと思います。つまり、遺伝体質、年齢、食事、ストレス等それらのいかんによって腸内細菌は変わり、その結果、老化や健康に関係してきます。
それではなるべく悪い腸内細菌にしないようにするにはどうしたらいいのかについてお話したいと思います。
最もいけないことは便秘だと思います。便秘をすると腸内にはどうしても悪い物質がたまります。したがって、たまったものはなるべく早く排泄することです。ちなみに、私達が食べ物を食べてその食べたものが大腸に達するまでの時間は大体五時間から七時間です。ですからあまり長くため込まないことです。そうかといって下痢をしてしまっては栄養が摂れませんが、便秘をしないためにどうしたらいいかといいますと、食物繊維を摂ることです。イギリスの医学者がかつての植民地アフリカへ行って診療してみてアフリカの原住民に大腸ガンとか高血圧、あるいは心臓病が少ないことに気がつきました。調べてみると、アフリカでは白い粉を食べないで皮ごとひいてパンのようなものをつくって食べている。つまり食物繊維を摂らないとこの文明病、現代病あるいは西欧病といわれる病気にかかることがわかったのです。いま日本人が成人病として悩んでいる病気の大部分がそれです。便秘、虫垂炎、痔、大腸憩室症、大腸ガン、潰瘍性大腸炎、過敏性大腸炎、大腸ポリープ、クローン病、そしていまかなりの若い人が悩んでいる肥満、それから糖尿病、心臓病、静脈瘤、血栓症、胆石、リウマチ様関節炎、腎臓結石、脳卒中、高血圧、そして老人性骨粗鬆症、悪性貧血等、日本人が老人病と言っているものがほとんど食物繊維を摂らないことによって起こるということがわかってきたわけです。このことから、食物繊維を多く摂れということが特に欧米で言われるようになりました。いまから十年ぐらい前のことだったと思います。
食物繊維は、皆さん菜っぱの筋をすぐ思い浮かべるかもしれませんが、それだけではなくて、コンニャクの粉など、水に溶けてドロドロになるマンナンやムチン等も含めております。そういう食べ物は穀類、特に雑穀にたくさんあります。それから豆類、サツマイモ、ジャガイモ、サトイモ等の芋類をはじめ野菜、果物、海藻(海藻には食物繊維だけでなくてミネラルが多く含まれています)、そしてキノコ類などです。食物繊維の効果といいますと、私は大体三つを考えております。一つは、腸内のビフィズス菌等を含めて有用菌がふえてくる。そしてその結果は有害菌が抑えられ、大腸ガンの予防に役立つのです。またビフィズス菌等は、乳酸と酢酸をつくりますと腸の運動を刺激し、便秘を予防することができます。食物繊維は消化することができない多糖体ですから、そのまま大腸に達します。そして消化することができませんから腸内でかさを増して、水分を吸い込み、これが一緒になり糞便をつくります。また、水をため込むと栄養分もため込んでしまいます。日本人はかつて戦争中は栄養が不足していて全部吸収しなくてはいけない時代だったのですが、いまは少しぜいたくになり、食べ物はたらふく食べています。ですからそのまま全部吸収したら太ってしまいます。したがって栄養分は少し捨てた方がいいのです。そのときにこの食物繊維が役立つわけです。そして栄養分をため込むとき糖質をため込んで糖の吸収を抑えます。だから、同じ糖を食べても、食物繊維を摂っていますと糖尿病になりにくく、糖尿病の予防になるということになります。また、便がかたくなりませんから、盲腸炎とか憩室症を防ぐことができます。さらにもう一ついいのは、水に溶ける食物繊維のムチンとか、サトイモのネバネバの成分のヘミセルローズ、あるいは海藻などは積極的にコレステロールをため込んで捨ててしまう働きがあります。コレステロールを捨ててくれますから心臓病にかかりにくくなります。現在の日本人の胆石はコレステロールが核になって胆石が大きくなるのですが、それを予防することができると言われています。こう考えますと、食物繊維はすべていいことばかりのように思われます。これは少し栄養過剰の現在のわたくしたち日本人にとってはきわめていいものだと思います。
かつては蛋白、脂肪、炭水化物、それからミネラルとビタミン、これを五大栄養素と言いましたが、いまはこの食物繊維が第六の栄養素と言われるようになったわけです。栄養素と言われますが、カロリーにもなりませんし、体に入って調節作用もないのですけれども、栄養学的にきわめて重要な成分になってきて、そしてこれを摂りなさいということが日本でも、あるいはアメリカでも奨励されております。日本でもだんだん食生活は欧米化されてきました。欧米化してくればくるほど食物繊維を多く摂る必要があります。それが欧米食によって腸の中でできる毒物を積極的に排泄する役目を果たしているのだというふうに考えていただいたらいいかと思います。
十五年ぐらい前から食物繊維の効用が言われたわけでございますが、私はこの食物繊維は本当に大切なのかということを腸内細菌の観点から調べてみたことがございます。新宿から甲府に行く中央線に上野原という駅があり、その北の方に棡原
そこでいただいた便を培養してみますと、先ほど言いましたように、若い人の腸内細菌は、ビフィズス菌が一〇%から二〇%ぐらいですが、それより少しは減っていますが、それでもビフィズス菌は五%から一〇%ぐらいみんないるのです。それから悪い菌の代表ウェルシュ菌、これが若い人並みで、二人に一人しか検出されません。それに対して東京都の老人病研究所の老人は十人中八人にウェルシュ菌がふえています。そして糞便のpHを見ますと、私達のはどんなに努力しても六・〇ぐらいですが、五・八ぐらいの人が多い。これを見て、遺伝的な体質もあるかもしれませんが、やはりこのような食生活がおそらく腸内細菌をよくして、長生きに貢献しているのではないかと私は実感したわけです。
長生きするためには心臓病とコレステロールというのは切っても切れない関係があります。これにも食物繊維を摂ることが大事だと思います。食物繊維はコレステロールをどんどん排泄してくれる。それからもう一つは骨粗鬆症、これは骨がグスグスになって折れやすくなる老人の典型的病気で、特に日本人に多いと言われております。なぜかといいますと、カルシウムの摂りかたが少ないからです。昔から日本ではカルシウムを摂るなら小魚を食べなさいとよく言うのですが、小魚よりは牛乳に含まれているカルシウムがきわめて吸収がよくて利用されやすいということで、ぜひ乳製品、特に牛乳あるいはヨーグルトをお摂りになると、カルシウムが多く摂れて、しかも骨がグスグスにならないで健康で長生きできるのではないかと思います。
日本人は、昔から牛乳はあまり摂っていないのですが、欧米の人は牛乳を多く摂っています。それに比例してカルシウムの摂り方は欧米人の方がずっと多いことがわかっています。また、食塩は脳卒中あるいは脳出血、高血圧にきわめて悪いということが、はっきりしていますから、なるべく少ない方がいい。しかし、味としてはある程度はほしいわけですが、できれば八グラム以下に抑えること。八グラムとはどのくらいかの目安に一つお話しておきたいのは、ラーメンです。ラーメンを召し上がると大体食塩が五―六グラムぐらい摂れてしまう。ですから、ラーメンを召し上がったら汁はなるべく残すことです。そして食塩は大体十グラム以下に抑えて高血圧にならないようにする。高血圧は動脈硬化、あるいは脳出血にもよくないですから、そんな工夫をされるといいと思います。ちなみに、東北は脳出血が多いと言いますが、大体秋田、山形では二十七―八グラム摂っております。日本の南部では十七グラムですが、これでもまだ多過ぎます。アメリカでは大体十グラムぐらい、またエスキモーはほとんど摂らないで魚などで済ましているのですが、それは四グラムくらいしか摂っていないので高血圧がないと言われています。このことでも高血圧と食塩とがいかに関係するかということがおわかりいただけると思います。
それでは今日私がお話したことをどうやって実行に移すかという秘訣をお話したいと思います。まず便秘がいけないので、毎朝排便すること。その時の便の色と形と硬さとにおいと量、この五点で毎日自分の健康状態がいいか、悪いかを観察されたらいいと思います。色は黄色っぽい方がいい。と言いますのは、胆汁の色素によって色が変わりますから、そうすると黄土色がいい。赤ちゃんのような黄色い便はちょっと出ませんが、黄土色の便を出すこと、茶色い便というときは、繊維が足らないか、蛋白が多過ぎるのか、あるいは脂肪が多過ぎるのではないか。特に脂肪が多くなりますと、胆汁がどんどん出てきますから茶色くなります。ですからなるべく黄色っぽい便を出すこと。それから、硬からず、軟らかからず、これはもちろんのことです。そして、その硬からず、軟らかからずの形はバナナ状の便になるわけです。コロコロ状、これは若い人、若い女性などがストレスを受けますと、兎糞と言ってコロコロと兎の便のようになります。これは何か自分がストレスを受けて悩みごとがあるときの便です。食物繊維が足らないときはコチコチ状の便になります。またお年寄りになりますと直腸型の便秘になります。どうしても直腸にたまってしまいますが、これも食事によって治すことができます。それから半練状というのは健康にとってそれほど悪いものではないのですけれども、あまり気持ちのいいものではありませんから、なるべくバナナ状にした方がいい。泥状とか水状、これは下痢ですから、何かおかしい、何とかして治さなくちゃいけないというふうにお考えになって、なるべく理想的なバナナ状の便、そして量は多い方がいいのです。日本人の便は昔から多いと言われております。便というのは民族によってずいぶん量が違うようで、統計的に言いますと、アメリカ人が大体一日に六十グラムから九十グラム、バナナの大体半本ぐらいしか出ないのですが、それではよくないと思います。年を取りますと少し少な目の方がいいかもしれませんが、若い人は大体一日にバナナ二、三本排便するのが目安です。それが大体百五十グラムから二百五十グラムの間ですから、年をとりましても、まあ百五十グラムぐらいは何とかして出すように工夫されるといいと思います。そして、もともと糞便は臭いものですけれども、腐敗産物の少ない便となりますと、においは少ないわけですから、あまりにおいの強くない便、このように色と形とにおいと硬さと量、これで自己診断していただくといいと思います。黒い便はよくないといわれておりますが、ノリなんか食べますと黒っぽくなりますし、ホウレンソウをたくさん食べますとやはり黒っぽくなりますから、これは昨日食べた食べ物で黒っぽいのだなというふうにお考えになればいいと思います。本当に黒いような便ですと、胃潰瘍か何かがあって出血して黒っぽくなっているのです。そのときには気をつけられて医者に診てもらったらいいと思います。
最後に、〝健康長寿のための八カ条〟として健康で長生きするためには成人病にならないことです。成人病にならないためには、動脈硬化もなるべく遠のけること。それには、第一に考えなくてはいけないのが食生活、そして心の問題、それから運動、この三つが重要だと思います。食生活では日本食にも欧米型の食事にも偏らない中庸がいいのではないかと思います。蛋白や、脂肪が少なくても脳卒中型になります。多過ぎると大腸ガン・乳ガン型になりますから、ちょうどその中間ぐらいがいい。数量的に言うと、牛肉だと百グラムから百五十グラムぐらいに抑えておくのがいいのではないかと思います。それからバランスのいい組み合わせ。そして自分は少し太る体質だと思ったら糖質を避ける、それから運動不足だと思ったらカロリーを抑えること、そして絶えず一週間に一度ぐらいはヘルスメーターに乗られ、自分の身長(cm)から一〇〇(cm)を引いて、それからさらに一割引いたキログラム、これが理想体重です。そのぐらいを努力目標にして、体重を維持していくこと。そしてそれにいま申しましたような便が出るように工夫して食物繊維を多く摂ること。それはなるべく食生活からお摂りになった方がいいと思います。しかし、食生活だけではなかなか摂りにくく、不足する面があります。その不足する分は、健康食品コーナーには、いまいろいろなものが売られておりますから、その中でいいものを選んで摂られること。食塩はなるべく摂らないこと、そしてビタミンCとE。これは体の中にできる悪い物質を無毒化、不活性化してくれますので、ぜひ積極的にお摂りになった方がいいと思います。食物繊維を一日大体二十グラムぐらいはお摂りになるといいと思います。二十グラムというとかなり努力しなくてはなりません。しかし、努力して摂ろうとすると食べ過ぎになりますから、それを健康食品として何かから摂られるのも一つの方法かもしれません。それから今日はあまりヨーグルトのことを申しませんでしたけれども、ヨーグルトというのは非常にいい食べ物です。これは何か別のいいものが入っているのですが、よくわかっておりません。しかし、ヨーグルトを食べますと寿命を長くします。これについては私のところで動物実験で証明しましたけれども、ネズミでも寿命が一〇%ぐらい長くなりました。また糞便の色がきれいになり、においも少なくなります。そして、できればビフィズス菌の入ったヨーグルトがいいのですが、これは限られてしまいます。ビフィズス菌は、特にお年寄りになりますと、少なくなったり、あるいはなくなっている方がいますから、そういう方は積極的に摂られた方がいいのではないかと思います。いまいくつもビフィズス菌の製品が出ておりますが、こんなに買えるのは日本だけで、外国はまだ全然ビフィズス菌というものはご存じないのです。これは本当に結構なことだと思います。ビフィズス菌を実際に摂ることによって私達の寿命は長くなると思います。それから疲労とかストレスは禁物です。なるべく睡眠とか休養とか楽しみによってそれは早く解消すること。疲労とかストレスがありますと腸内細菌のバランスが悪くなって悪いものがつくられますから、なるべくストレスを受けないようにする。一番いいのは、悟りの境地を開くことが最もストレスや疲労を受けない、いい方法ではないかと思います。
もう一つ重要なのは生きがいだと思います。自分は人類のために尽くし、これだけ生き抜いたのだという自負心を持って、もっと生きなくてはいけないと、このことが生きる力になって、活力になると思います。つまり、悟って生きがいを感ずること、これがいいのではないでしょうか。そうするとストレスなどは飛んでしまいます。そして適度な運動、これも適度な、でいいのですが、まず歩くことです。それから入浴。歩くことが少ない人は特に入浴し体をきれいにし、血行をよくします。それが腸の運動にも関係して、便秘を解消することにもなります。タバコはなるべく吸わないこと。お酒はほどほどに飲むこと。適量というのは人によって違いますので、ぜひ自分の適量を見つけて、ストレス解消に、あるいは楽しむ程度には、お飲みになる方がむしろいいかもしれません。
バランスのいい食生活で、しかもビタミンCとE、それから食物繊維、ヨーグルト、あるいはビフィズス菌、こういうものはなるべく多めに摂る、そして心は豊かに生きがいのある生活をすることによって健康で、寿命ももっと長くなるのではないかと思います。ある人は日本人はもうだんだん長寿から短命に変わると言っておりますけれども、私は日本人のように知的意欲の強い国民はおそらくもっと世界一を守り続けるのではないかと思います。今日のお話が皆さんの健康と長寿に役立つことができれば幸いだと思います。
ご清聴ありがとうございました。
(東京大学教授・理化学研究所主任研究員・東大・農博・昭28)
(本稿は平成元年10月20日午餐会における講演の要旨であります)